
Ghostly Shadows at War
What Perception Disorders Reveal About Consciousness – Part 1

Some blind patients can catch a ball or dodge an oncoming cyclist, despite not seeing either. Meanwhile, some sighted patients with perfect eyesight cannot see motion at all, and would be struck by both. These cases challenged how vision was understood and still raise questions about consciousness itself.

We have spoken about LM, a woman who, despite having perfect sight, had lost the ability to see motion and could see the world only as a series of still images. If you have not read that, you can catch up here. But what if the opposite is also possible? Can someone see an object move while not being able to see the object itself? If so, what do they see?
Vision is associated with the primary visual cortex, also called V1, at the back of the brain. In 1905, anatomist Paul Flechsig wrote that V1 was “the only entering place of the visual radiation into the organ of psyche”, and by that he meant that V1 is the only place where visual information enters consciousness.
WW1 and ghostly motion
British neurologist Captain George Riddoch served as an officer in the Royal Army Medical Corps during the First World War. He treated soldiers who were blind due to gunshot wounds to the primary visual cortex (V1). This was a common wound as the helmets left the back of the head exposed.

Riddoch mapped the visual fields of each patient to define the size and shape of their blind field. To his surprise, some patients reported that sometimes they could see “something moving” inside their blind fields. For example, one soldier could see the motion of the feet of the people walking by, but nothing above the ankles. Another said he could see the motion of the spoon when he was being fed. Even though they were convinced they could see something, what they saw was “so vague and shadowy” (Riddoch, 1917).” The soldiers found it difficult to describe what they saw:
“The ‘moving things’ had no form, and the nearest approach to colour that he could attribute to them was a shadowy grey” (Riddoch, 1917).
Could it be that motion was not merely a component of vision but a separate feature of visual perception altogether?
What is Riddoch syndrome?
Riddoch syndrome is a rare neurological disorder in which people blind from damage to the primary visual cortex can still consciously perceive movement in their blind field, while remaining unable to see shape or colour. Depending on how much of the cortex is damaged, the blind field may cover the whole visual field or a portion of it.
What Riddoch saw
Riddoch’s 1917 paper challenged the accepted view of vision. At the time, V1 was considered the only gateway to conscious sight, and vision itself was thought of as a single sense, divided into light, colour, and form. Motion was not even in the picture. Riddoch’s observations suggested otherwise. By suggesting that patients could see motion without having a functioning V1, he was proposing that motion has its own neural basis outside the brain’s “visual centre.” Riddoch went further and suggested that vision was not unitary but made up of distinct perceptions and that they could fail independently.
Not surprisingly, his idea did not catch on. Not only because it was a heretic notion at the time, but also because there was another plausible explanation.
Same front, same side, different interpretation
At the same time Riddoch was studying this phenomenon, same war, same side, same city, Irish neurologist Gordon Holmes was also finding patients who could see movement inside their blind fields.
Holmes was stationed at No.13 Stationary Hospital in Boulogne, barely 2 km from the Empire Hospital, where Riddoch worked. But, while Riddoch was treating a handful of wounded officers in a repurposed private clinic, Holmes was treating soldiers on a ratio of 90 patients to 1 doctor (secondary account, but many, in any case).

Under those conditions, Holmes mapped the visual fields of around 400 soldiers who had damage to their visual area due to gunshot wounds. But if he found the same phenomenon, why did he disagree with Riddoch? Well… asking the right question is everything. Both men were mapping visual charts by asking patients to report when they saw a disk on a screen, and then plotting that on their chart (see image above). But Riddoch did not ask his patients to report only when they saw the object, but also when they saw it move. By comparing the answers, he could show that soldiers were detecting movement in the same area where they were blind.
Holmes dismissed Riddoch’s findings, suggesting that this “shadowy” visions could be explained by small patches of V1 that had survived. Holmes’ opinion carried more weight, as he was already a recognised neurologist and Riddoch’s former teacher. Riddoch’s theory was dismissed.
What evidence was missing?
To challenge Holmes’ rebuttal, Riddoch needed to show a double dissociation. It is not enough to see motion without shape and colour; he would also need the reverse: shape and colour without motion. If only such a case had been reported… (and if you have read this article, you are probably smiling…). However, that took a long time to come.
Riddoch is back, baby!
In 1983, Zihl and colleagues published the case of patient LM, a woman with clear vision of colour and form but no motion. She had intact V1 but damage to area V5 (more on V5 soon). This was the missing puzzle piece that finally cemented V5 as an area dedicated to visual motion, confirming what Riddoch had proposed 66 years earlier.
[Riddoch died in 1947, aged 58]
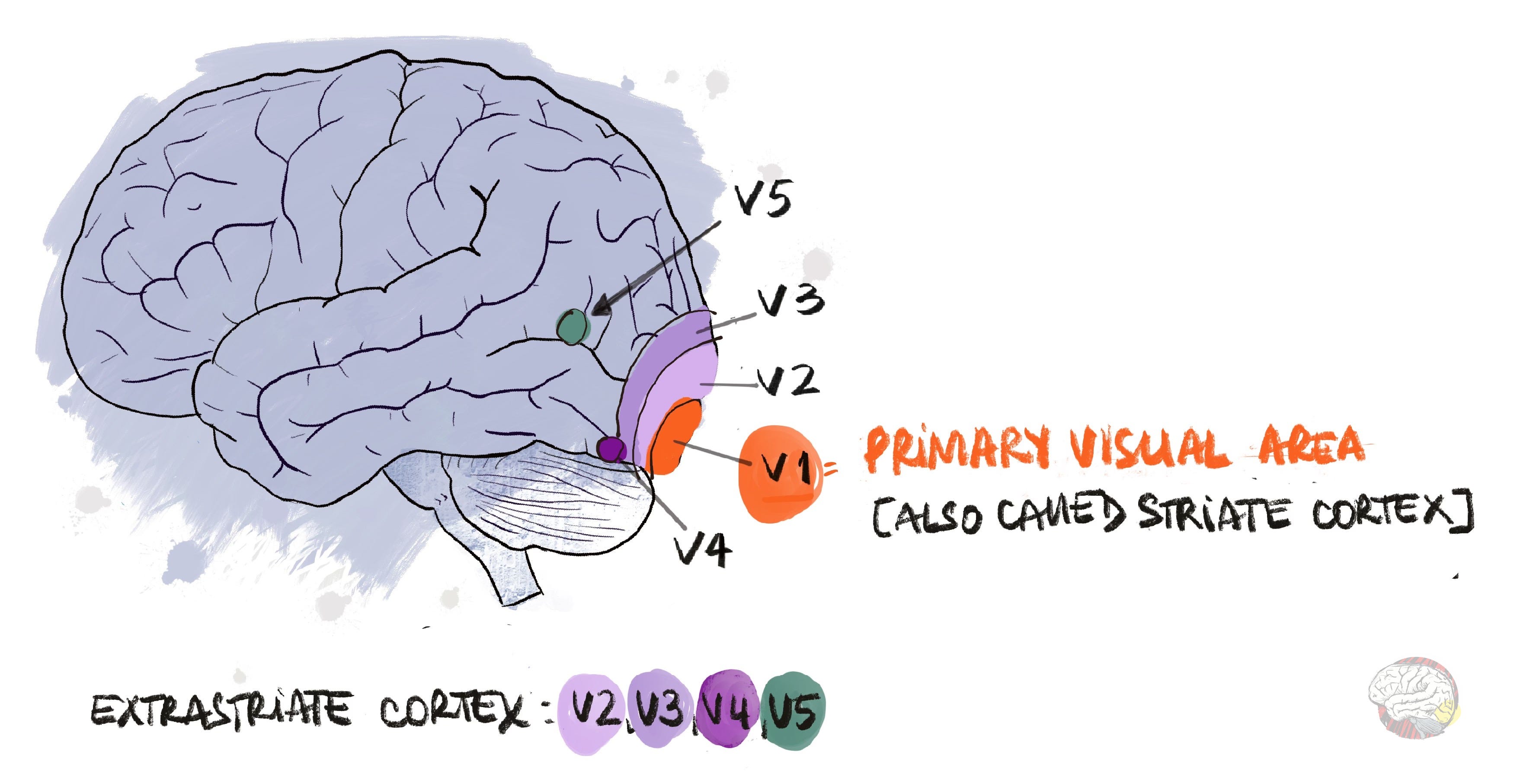
By the time LM appeared, the old idea that V1 was the only visual processing area was already beginning to crack. Research on animals in the 1960s–70s had already uncovered new visual areas beyond the primary visual cortex: V2 and V3 for integrating form, V4 for colour, and V5 for motion. Animal brains are useful models for mapping pathways, but they are not identical to ours, so findings always need to be confirmed in humans. That is where LM fits in: her case proved that perceiving shape and colour can be separated from perceiving motion, and confirmed what Riddoch had anticipated: that different visual features could fail independently.
If you are curious about the chronology, see here:
1960s – Hubel & Wiesel: Single-unit recordings in cat and monkey visual cortex; discovered orientation-selective cells and hierarchical processing. This set the stage for mapping extrastriate areas (outside of V1).
Early 1970s – V2 and V3: First outlined by Semir Zeki; mapped in detail by David Van Essen and John Allman.
1973 – V4 (colour): Identified by Zeki as a colour-selective region; later expanded by Margaret Livingstone & David Hubel in the 1980s.
1970s – V5 / MT (motion): Described by Zeki; its functional role confirmed in the 1980s by William Newsome, William Movshon, David Albright, and Ralph Siegel, who showed MT activity was tightly linked to motion perception.
I have left the references out to keep this timeline light. All of these findings come from classic visual neuroscience papers.
The science: understanding the visual brain
Vision is not processed in a single location in the brain. Instead, it is distributed across a network of specialised regions, and information about each feature (colour, shape, motion) is processed as separate streams and then integrated into what we experience as a seamless image.
Where Riddoch patients see motion without shape or colour,
those with motion blindness see shape and colour without motion.
The Role of Area V5 in Motion Vision
Riddoch patients could see motion because the area V5 in their brain (also called V5/MT) was intact. V5 is the brain’s motion centre. When it is damaged, as in LM, the fluidity of the world itself disappears. Cars appear to jump from one spot to another, people seem to teleport across rooms, and lips move out of sync with voices. Motion is simply gone. This condition, known as akinetopsia or motion blindness, is the mirror opposite of Riddoch syndrome.
Where Riddoch syndrome preserves motion vision without conscious form, akinetopsia (motion blindness) removes motion from a world otherwise fully visible. The two conditions represent opposite disruptions in the visual pathway and underscore the modular nature of perception.
Area V5
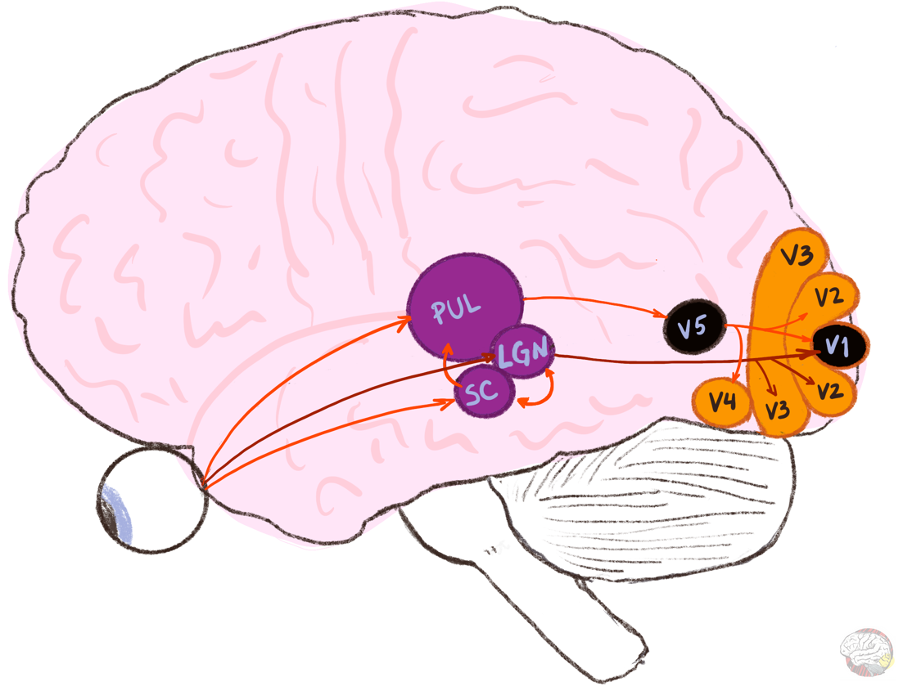
In normal vision, V5 receives its main input from V1 (Cragg, 1969; Zeki, 1969, 1971). But if Riddoch patients do not have a working V1, how does V5 still get the information?
See? You are already a step ahead of me. There are indeed alternate pathways to V5.
Alternative routes
Over the last few decades, studies in monkeys showed that several subcortical pathways bypass V1 and project directly to V5:
The superior colliculus —> It detects sudden movement and drives the eyes and head towards it. It also processes unconscious visual information. It connects to V5 via the pulvinar.
The pulvinar (in the thalamus) —> It integrates information from several visual areas and acts as a“gatekeeper,” prioritising signals and linking them to attention and emotion. It also connects to the amygdala.
Lateral geniculate nucleus (LGN, also in the thalamus) —> It is the retina's main relay to the cortex. Most of its visual output goes to V1, but a small shortcut goes straight to V5.
These circuits do not carry fine detail. The signal is fast and coarse, but it offers a survival advantage: it provides a fast lane that warns you if something is hurling towards you.
Modern times – neuroimaging evidence
Until now, all evidence was based on behavioural tests. Barbur and colleagues (1993) were the first to show the brain activity that matched what Riddoch had proposed. They used PET to study patient GY, a 36-year-old patient who was blind in half of his visual field (called hemianopia) due to a car accident at the age of 8. This provides neuroimaging evidence for the first time that V5 could be activated even when there is no activity in V1, and that this activation corresponded with conscious perception of visual movement.
This tells us three important things:
V5 receives information even when V1 is not functioning
Activity in V5 is enough to produce a conscious experience of movement
V1 is not the sole seat of vision, and is not the only place where visual information enters consciousness
Since 1993, fMRI and MEG have taken this much further. Over three decades of work have refined our understanding of how V5, subcortical inputs, and consciousness fit together.
Why this matters
Conscious awareness had long been linked to the primary visual cortex. But Riddoch shows that motion, at least, can breach consciousness without V1. This forces us to confront the fluid boundaries between seeing and not seeing, knowing and not knowing. This calls for a more nuanced model of how awareness emerges from the brain.
Consciousness in fragments
We know now that vision is not unitary but rather different features such as colour, shape, motion, integrated back together. It is not an “all or nothing” phenomenon, and patients with different lesions can see different aspects of the world. But what about consciousness? Is consciousness all or nothing? Or is it a patchwork of separate fragments of awareness?
Coming next…
Riddoch’s patients could see ghostly shadows move in their blind fields. LM could see the world frozen in still frames. But what if I told you that some blind patients can avoid obstacles without knowing they are doing it? Basically, they can see, but they are not aware that they can see. These cases gave rise to the concept of blindsight (vision without awareness) and forced science to rethink not just how we see, but what it means to be conscious.
Continue to Part 2 here.
Acknowledgement:
A warm thank you to Dr. Ahmad Beyh for taking the time to read this article (Parts 1 and 2) and share thoughtful feedback.
It takes a lot of work and time to write these articles. If you find value in what you just read and want to support my work, you can buy me a coffee.
In any case, if you got this far, please like and restack, and feel free to drop any questions in the comments.
References:
Barbur, J. L., Watson, J. D., Frackowiak, R. S., & Zeki, S. (1993). Conscious visual perception without VI. Brain, 116(6), 1293-1302.
Cragg, B. G. (1969). The topography of the afferent projections in the circumstriate visual cortex of the monkey studied by the Nauta method. Vision research, 9(7), 733-747.
Campbell, A. W. (1905). Histological studies on the localisation of cerebral function. University Press.
McFadyen, J. (2019). Investigating the subcortical route to the amygdala across species and in disordered fear responses. Journal of Experimental Neuroscience, 13, 1179069519846445.
Riddoch, G. (1917). Dissociation of visual perceptions due to occipital injuries, with especial reference to appreciation of movement. Brain, 40(1), 15-57.
Zeki, S. M. (1969). Representation of central visual fields in prestriate cortex of monkey. Brain research, 14(2), 271-291.
Zeki, S. M. (1971). Cortical projections from two prestriate areas in the monkey. Brain research, 34(1), 19-35.
Zihl, J., Von Cramon, D., & Mai, N. (1983). Selective disturbance of movement vision after bilateral brain damage. Brain, 106(2), 313-340.
Fascinating, thanks 🙏
What an interesting read. I almost expected it to conclude with an “I found Bigfoot” revelation, but the depth and clarity made it far more rewarding. Although my background is not in neuroscience, I always enjoy learning something new. As I read, I was reminded of the so called “phantom leaf effect,” where researchers photographed a leaf, cut part of it away, and then captured an image that still showed the outline of the missing section. Later studies challenged and largely debunked the phenomenon, but the idea remains intriguing.
It raises the question of whether, beyond the classic V1 to MT pathway, other regions might contribute in unexpected ways. For example, could the somatosensory cortex, typically associated with touch and bodily awareness, play a role in perceiving subtle shifts in movement or energy? Has there been any evidence suggesting that we can directly sense atoms or energy? If we and everything around us are composed of molecules and atoms, then it seems at least possible that the brain might be able to register these fundamental movements at some level, even if only indirectly.